

John Hawks & “Principles of Phys Anth” students at Dreamland BBQ (the original, natch)
University of Wisconsin-Madison paleoanthropologist John Hawks was UA’s second ALLELE lecturer of the season. Hawks was trained at the University of Michigan in anthropology by the famous Milford Wolpoff (he of multiregionalism infamy) & completed a postdoc in evolutionary genetics under Henry Harpending at the University of Utah. Hawks also has one of the most visited science blogs in the world & gave us a hat tip post prior to his visit (& now I am blogging about his blogging about us). His main ALLELE lecture was entitled “Neandertime! What Ancient Genomes Can Tell Us About Our Past & Present” (which should soon be up on iTunesU for you to enjoy in its entirety here). Dr. Hawks arrived the night before, in time to be taken out for Dreamland BBQ by our grad students, who have been blogging about his blogging in anticipation (see “Hello Archaeology, We Meet Again,” “John Hawks: Superhero,” “What the Hawks is Up?,” “John Hawks, Daniel Lieberman, & ‘The Stalin Age Diet’,” “Plaque Attack: How Food Fought Back,” & “John Hawks Week at UA“). He also gave a talk to the Anthropology Department entitled “Human Evolution in the Holocene,” let himself be trotted thru lots of meetings & meals with faculty & students across the University, including a finale of pizza & CAKE!

John Hawks & CAKE (we stole this idea from SUNY New Paltz, home of the original EvoS cake). Photo by Max Stein.
The overall takeaway is that John Hawks is a GREAT speaker, as he is extremely affable & blows minds for a living. As hosts (the Evolution Working Group), we’ve learned to look for speakers who will both teach us AND be accessible to students & an educated public, & John Hawks can do just that. Some topics in evolution, it turns out, are sexier than others, &, apparently, Neandertals, big strapping fellows that they were, still have it. I will admit that our Anthropology-sponsored speakers aren’t known for pulling down the audience numbers that, say, Biology-sponsored ones do (perhaps because Bio simply have larger enrollments in core courses they can require/bribe to attend?), & we were worried that a speaker during the last week of class would be overshadowed; but the turnout was nothing short of impressive for both talks. And it didn’t hurt that John could seriously talk BBQ & football in Bama!

Neandertals have still got it! Look at the great turnout for the last day of the semester (photo by Max Stein).
I am blown away by a few key points made in his talks & in his writings over the past several years. I think the biggest mind-effer from the ALLELE talk is that the Denisovans represent a new species of our genus that we never suspected exist. Of course many Homo species have come & gone over the years, as fossil remains have been reclassified (see Erin Wayman’s “Hominid Hunting” Smithsonian blog posts “Four Homo species you’ve never heard of I” & “II“), but genome sequencing–as has been done with the Denisovan remains, Neandertals, & several other hominid species–enables us to calculate degrees of relatedness & antiquity with accuracy previously unrealized. Couple this with the Homo floresiensis finds from a 8 years ago (word has it that we can now no longer refer to this as “The Hobbit”), & what this implies is that there are possibly many more species that co-existed & combined with our ancestors than we ever considered.
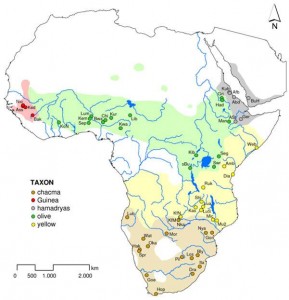
Savanna baboon parapatric distribution in Africa (Source: Zinner et al 2009).
On that note, the genetic admixture of these varied species is low, indicating that conversations about Neandertals & H. sapiens interbreeding are overblown. But what it does not say is that it never happened. Hawks suggests that the distribution being uncovered may be the result of parapatric speciation of the Homo genus in antiquity like that observed today among savanna baboons. There are a number of savanna baboon species distributed across Africa that look very similar but largely do not interbreed because movement between groups is extremely dangerous & is likely to get individuals killed. But limited hybridization does occur rarely along the species borders where familiarity can develop. It may be that Homo species behaved in similar ways, hybridizing with one another rarely near population borders, a scenario which irretrievably muddies the lumping/splitting heuristic of paleoanthropology that we diagram for students. For instance, the Denisovans & Neandertals, despite this overlapping distribution, are no more closely related than Kalahari Bushmen & Amazonian Indians. They do share some attributes with all hominids at northern latitudes. Both Neandertals & Denisovans had mutations in pigmentation genes that probably lightened them in ways different than similar genes in H. sapiens.

John Hawks, Denisovan weatherman (Photo by Max Stein).
What is surprising about the Denisovans is that there is evidence of genetic relatedness in living populations today but not in Asia where one would expect it. This evidence is found in Austronesian populations & seems to have been completely displaced from Asia. Additionally, some Denisovan genes are found at higher rates than for that of the overall genome, suggesting these genes were immunologically important. What I had been waiting for is how all of a sudden, we have a new species of Homo making a splash in the news, as Denisovans did a few years ago, though seemingly little has been written about their fossil remains since. This is because, as it turns out, all of this comes from a bone sample taken from one girl’s finger bone found in Denisova Cave in Siberia, which they had initially suspected was either Neandertal or Homo sapiens, both of which seem to have occupied the area at different periods. To discover that the area was shared by others was a shock. Perhaps this was a border region for the various populations, but like Georgia in the Middle East.

Dr. Evil? Or the whole Denisova genome from one girl’s finger bone? (Photo by Max Stein)
On the other hand, Neandertal genomes display previously unappreciated diversity. What was believed to be a largely European species, overlapping with H. sapiens, now appears to have been largely based in Asia. European Neandertal DNA from after 50kya was less diverse than that found in Asia or in Europe before 50kya, leading to the conclusion that Europe was more like a backwater for Neandertals. A few migration events seem to have take place from Asia to Europe by groups widely separated in time & genetic relatedness. They were genetically only distantly related to Asian Neandertals & earlier Neandertals in Europe. We know more about Neandertals from genomic analysis of aspirin-sized samples of old rotten bone, otherwise useless to anatomists, than any of the fossils that have been found. Still, the patterns of Neandertal genetics still unclear. For instance, a dopamine gene variant may be implicated in H. sapiens movement out of Africa (going where no man has gone before, Hawks suggests, like “Captain Kirk”; thus, I am dubbing it the “Captain Kirk” dopamine polymorphism). Hawks likens this to the vanguard of the can toad invasion in Australia, which were found to have longer legs than the toads behind the front wave. Neandertals were a thrill-seeking people (jumping on bison with spears for dinner–2/3 were dead by age 30, whereas H. sapiens were using distance weapons) & carried the Captain Kirk gene, but the distribution in their population is unclear.
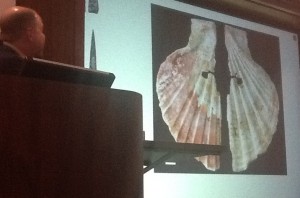
Neandertals used red ochre to decorate shells
Neandertals have an undeserved reputation for being unsophisticated. “NeanderTHals” means “stupid” in our culture; “NeanderTal” implies more sophistication, like saying “organic produce.” Neandertals were not stupid cavemen, but they were chauvenists–they tended to bury only their men! Neanderthals used to be the “missing link” between ancestors & Homo stupidous (seriously). There is evidence they systematically removed dark raptor feathers to adorn themselves (“Neandertal goth”!). They also systematically gathered pigmented shells & colored the other side of them with red ochre in the same manner. Though evidence of fire use appears only inconsistently in relation to Neandertal living floors, evidence of processed starch grain in dental calculus (the gook that built up on their teeth) indicates they were cooking in a moist way. The irony is they had no cooking vessels, so how were they cooking this, Hawks queries? Banana leaf tamales? Haggis? Though they did make pigment cups from the tips of stalagmites. Language is a keystone aspect of H. sapiens culture. We can’t verify that Neandertals could talk, but they had audio channels similar to humans & different than chimps.
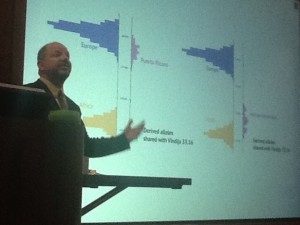
More Neandertal relatedness among Europeans (top-blue) & Puerto Ricans (top-pink) than Africans (bottom-yellow)
When comparing modern populations, we see 5% more relatedness to Neandertals outside of Africa than inside Africa & more relatedness among Asians than Europeans. This holds true among sub-populations of migrants from these regions as well. However, it appears that modern Europeans are different than those of just 5,000 years ago. Genomic analysis of Otzi, the frozen mummy of a Tyrolean man found in the Italian Alps in 1991, indicates that Europeans used to have a higher degree of relatedness to Neandertals. We don’t find people in Europe today who appear to have come from Otzi’s population genetically.
We don’t know why the Neandertal populations diminished in Asia or Europe & ultimately disappeared, but Hawks speculates that, as with extinction events today, H. sapiens may have introduced diseases to which Neandertals had less immunity. Hawks points to the plight of the red/gray squirrel as an example of how this may have happened (gray squirrels are recent immigrants & seem to have displaced the red squirrels–remember Squirrel Nutkin?–by introducing a parapoxvirus to which the red squirrels slowly succumbed).

Dr. Hawks talks with students over pizza & cake
From his Anthropology talk, my takeaway was that evolution has sped up in humans over the past 40,000 years, not slowed down as previously believed. Population geneticists like Hawks have written a lot on the importance of population size over the past several years, & I have simply failed to notice it (here is a recent post by John discussing another article that addresses this), but it makes intuitive sense. The likelihood that any mutation, whether beneficial or not, will persist in a population is based on the population size. If we consider genetic drift, we know it is more effective in small populations, as allelic variation has a higher likelihood of drifting out & allowing genetic fixation to take place. The same is true when we consider our history with pathogens–virulent communicable diseases burn themselves out when population sizes are small after they kill off all the susceptibles but can persist if population sizes are large & distributed enough to enable them to spread slow enough for susceptibles to be replaced. The likelihood of a mutation remaining in a population is greater if that population is big & distributed enough for gene flow to be effective. This is the case not only of beneficial mutations but also of slightly deleterious ones. In fact, it may be beneficial in the long run that even negative variations be maintained, as they can be co-opted & turned into benefits when environments change.
Hawks suggests that population density was critical for the human cultural revolution of the Upper Paleolithic to happen, insinuating that it was both genetic (as Klein & Edgar have suggested [see Hawks review]) & gradual (as McBrearty & Brooks [2000] propose). Thus, if we have a way to measure population size, we have a means to measure natural selection. Given the tremendous growth of the human population over the past 8,000, we should have a flood of mutations, & many have been found. We know mutations for celiac disease & other disorders have arisen recently. There is evidence that human evolution has recently sped up. If we speculate based on the 500 found dating to the past 40,000 & calculate based on a constant mutation rate going back 2.5mya when humans & chimps diverged, there should be 750,000 separating the two species. However, there are only 1,200 separating us.
Lactase persistence is probably the most well-known example of recent human evolution. It was a mutation in only one person only 8,000 years ago (when the population is estimated to have been <5,000 people) & is dominant in 75% of Europeans today. None of the remains analyzed in Germany from 5,000 years ago have the lactase persistence gene, not because it’s not there but because it’s still rare. Another are hemoglobin gene polymorphisms, which have selection histories intertwined with malaria. Ironically, the HbE variant that appears in Southeast Asia conveys resistance to Plasmodium falciparum, the most virulent form of malaria in Africa. When Hawks asked Frank Livingstone (the anthropologist who synthesized a classic 4-field anthropological model to explain the selection for sickle cell in Africa because of cultural practices) why HbE never developed in Africa, Livingstone’s answer was, “because it didn’t.” That’s how evolution works. It is a random process.

Meeting with EvoSers & HBERGers (my lab)
Finally, Hawk’s blog essentially inspired me to start blogging & to inflict it on students, so I asked him tell students some of the benefits of academic blogging. One of the coolest points he made is that just a few hundred hits on one free blog post is actually more traffic than many museum & other outreach websites get on their whole site, though those sites cost them lots of money to build & maintain. This is a great argument for the utility of blogging as a means of dissemination & service to the discipline. Blogging can be slow to take off–first one attracts journalists who want an expert rather than do the research themselves, then one attracts other professionals, then the rest. Hawks started blogging when it was relatively new after struggling to get a website going. “At every stage, people said it was insane what I was doing, so I never tell students to do what I did.”

Pingback: 2012′s Cheap Thrills thru Evolution in Review | Welcome to the EvoS Consortium!